Lei Luo ![]() ,
Yingchao Hao,
Li Jia,
Wenxue Zhu
,
Yingchao Hao,
Li Jia,
Wenxue Zhu
For correspondence:- Lei Luo Email: 13623896431@139.com Tel:+13623896431
Received: 9 April 2015 Accepted: 3 January 2016 Published: 28 February 2016
Citation: Luo L, Hao Y, Jia L, Zhu W. Analysis of chlorogenic acid oxidation pathway in simulated enzymatic honeysuckle browning system by high performance liquid chromatography and mass spectrometry. Trop J Pharm Res 2016; 15(2):405-409 doi: 10.4314/tjpr.v15i2.25
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the pathways involved in the oxidation of chlorogenic acid (CA) and phenol metabolism in honeysuckle buds.
Methods: A model that mimics CA oxidation by honeysuckle polyphenol oxidase (PPO) by controlling the reaction temperature or reaction duration was employed, and the resulting products were analyzed by high-performance liquid chromatography followed by mass spectrometry (LC-MS).
Results: The pathway of CA oxidation by PPO in honeysuckle involves CA, hydroxyl - CA, CA-o-quinone derivatives, and CA-p-quinone derivatives.
Conclusion: CA oxidation and the enzymatic browning reaction significantly impact the quality of honeysuckle and other fresh agricultural products.
Introduction
Honeysuckle, which is the flower bud of the Caprifoliaceae plant (Caprifoliaceae lonicera L.), is an edible natural substance [1] with medicinal properties used in traditional Chinese herbal medicine. Because it possesses a variety of biological activities, honeysuckle is widely used in pharmaceuticals, beverages, health foods, and other products. One of the major bioactive components of honeysuckle is chlorogenic acid (CA).
Phenolic compounds in plant-based foods are important factors that contribute to enzymatic browning, a process that commonly occurs in fresh plant-based foods. There are many phenolics that induce browning [2], but CA is one of the most common. The widely accepted mechanisms that cause browning include mechanical damage (e.g., peeling, cutting, bruising, insect bites, or pulping) or abnormal environmental conditions (e.g., frostbite or heating) that destroy the redox balance in fruit or vegetable tissues and lead to the accumulation of oxidation products, color changes [3], and severe reductions in commercial and nutritional value. The water content of fresh honeysuckle is approximately 80 %, which renders the flower unsuitable for transport and storage. In order to ensure the quality of honeysuckle buds and to prevent the deterioration of color and the accumulation of bioactive components [4-6], immediate drying is required prior to preservation.
In a prior study, Hou [7] demonstrated that enzymatic oxidation of CA is the major cause of the browning reaction in honeysuckle buds. Additionally, Robards et al [8] revealed that differences in the generation of quinones induced by enzymatic and non-enzymatic browning are a result of the rate of enzymatic browning, which occurs much faster than non-enzymatic browning. Accordingly, the purpose of this study was to establish an enzymatic system of honeysuckle polyphenol oxidase (PPO) and to evaluate the CA oxidation pathway in honeysuckle.
Methods
Reagents
The analytical reagents used to prepare the PPO included analytical grade phosphoric acid, alcohol, phenylthiourea, ammonium sulfate, disodium hydrogen phosphate, and sodium dihydrogen phosphate. Chromatographically pure acetonitrile and methanol were used as the mobile phases for high-performance liquid chromatography (HPLC). Chlorogenic acid was purchased from the National Institutes and was used as the food and drug control (Beijing, China).
Plant materials
Fresh Jinfeng I honeysuckle (Caprifoliaceae lonicera L.) buds were harvested in Fengqiu County, China. The buds were picked once per month from July to September 2012. The plants were authenticated by Weiping Yin, president of the College of Chemical Industry and Pharmaceutical at Henan University of Science and Technology in June 2012.
Preparation of PPO from honeysuckle buds
Fresh honeysuckle buds were screened for pests and washed with distilled water. After drying, the juice from the honeysuckle buds was weighed and mixed with 0.05 mol/L phosphate buffer (pH 6.5) containing 4 % polyvinylpyrrolidone (PVP) at a 1: 4 (W/r) ratio to obtain honeysuckle bud homogenate. The homogenate was then extracted with methyl alcohol for 24 h at 4 oC, filtered at 4 oC, and centrifuged for 30 mins at 12,000 rpm. The supernatant was collected as the crude extract. To obtain 95 % saturation, the extract was centrifuged at 12,000 rpm for 30 min at 4 oC. The precipitate was dissolved in 5 mL of 10 mM phosphate buffer (pH 6.5) and dialyzed against the same buffer for 24 h at 4 oC [9-11]. The dialyzed sample was used in the experiments.
Preparation of enzymatically-oxidized products in the mimic system
In this study, CA was selected as the phenolic material standard. The pH of the reactions was adjusted to 6.83, and the concentration of the CA was 200 mg/L. The ratio of PPO to phenolic substances was 1:2. The enzymatic oxidation reactions were conducted at 0 oC for 0, 0.5, 1, 10, and 35 min (as the enzymatic oxidation system I of CA). Reactions were also performed for 1 h at 0, 20, 40, 60 and 80 oC (as the enzymatic oxidation system II of CA).
Preparation of non-enzymatic oxidation products in the mimic system
The non-enzymatic oxidation reaction was conducted at pH 6.83 in the presence of 200 mg/L CA for 1 h at 40 oC.
HPLC analysis of oxidation products in the mimic system
After completion of the mimic reaction, methanol was added to the reaction system to a final concentration of 50 %, and extraction was conducted in a water bath for 1.5 h at 70 oC. Prior to HPLC analysis, particulates were removed by filtering the extract through a 0.45 µm membrane. For analysis, 0.5 µL of the extract was injected into a BEH C18 analytical column (2.1 mm × 100 mm, 1.7 µm). The mobile phases consisted of acetonitrile (A) and 0.1 % formic acid (B). The extracts were eluted with the following gradient: 5 – 40 % A for 0–20 min and 40 – 90 % A for 20 – 25 min at a flow rate of 0.3 mL/min. The column temperature was 45 oC and the detection wavelength was 327 nm. CA was used as the internal standard. Under each reaction condition, the components of the oxidation products were compared and analyzed by mass spectrometry.
Analysis of significantly changed products in the mimic system
For analysis, 0.5 µL of the extract was injected into a BEH C18 analytical column (2.1 mm × 100 mm, 1.7 µm). The column temperature was 45 oC and the mobile phases consisted of acetonitrile (A) and 0.1 % formic acid (B). The extracts were eluted with the following gradient: 5–40 % A for 0–20 min and 40–90 % A in 20–25 min at a flow rate of 0.3 mL/min. All extracts were analyzed by electrospray ionization (ESI) by applying a 3.5 kV capillary voltage within the ion source. The cone voltage was 20/40 V, the ion source temperature was 100 oC, and the de-solution temperature was 400 oC. The flow rate of the de-solution gas was 500 L/h and the flow rate of the cone gas was 50 L/h. The MS survey scan range was 100–1500 m/z, the collision energy was 6 V, and the detector voltage was 1600 V.
Data analysis
The data resulting from the preparation of PPO were analyzed using Design-Expert® software (v. 8.0.6). P < 0.05 was considered statistically significant. The data from HPLC and mass spectrometry experiments were analyzed using Waters MassLynx™ (v. 4.1) MS software.
Results
The CA enzymatic oxidation process at various reaction times
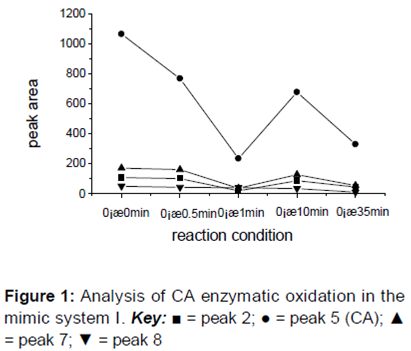
The results revealed the status of each of the reaction substances during the oxidation of CA by PPO at the initial time point (0 min). Due to the extended reaction duration, obvious changes were detected in the reaction substances over time. All of the chromatographic peaks were numbered, and the peak areas at different retention times were compared after omitting peaks with no apparent changes and generating a curve that described the change in peak areas ().
shows changes in the concentration of the reaction substances during the CA mimic enzymatic oxidation reaction at 0 oC after 35 min. Although the areas for peaks 2, 5, and 7 were larger at 10 min than the corresponding peak areas at 1 min, a gradual trend indicating the reduction in total reaction substances was observed over the course of the 35 min reaction.
Enzymatic oxidation of CA at various temperatures
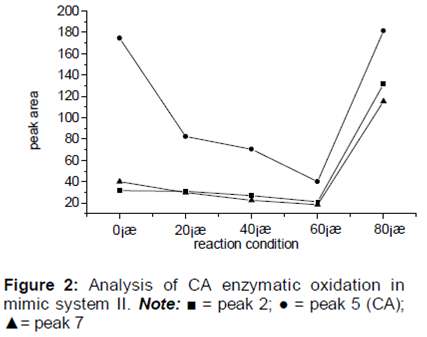
The results demonstrated the catalytic oxidation of CA in 1 h at 0, 20, 40, 60, and 80 oC. All of the chromatographic peaks were numbered and compared. A curve describing the obvious changes in the peak areas is shown in .
Exploration of the CA enzymatic oxidation pathway revealed that the PPO activity at low and high temperatures (0 and 80 oC, respectively) was inhibited, whereas the enzymatic oxidation reaction at 40 oC was the strongest and thus the analysis of the CA oxidation products generated at this temperature was more difficult (). In order to facilitate the control of CA oxidation products at each stage, the enzymatic oxidation of CA in the mimic system was held constant at 0 oC.
reveals comparable trends in the peak areas of the three substances represented by peaks 2, 5, and 7, which suggested that these substances were similar to CA and had a consistent reaction pathway in the reaction system. Following the HPLC analysis of the products of the CA enzymatic oxidation process after various reaction durations, the substances from peaks 2, 5, and 7 were believed to be phenolic acids.
Enzymatic oxidation pathway for CA
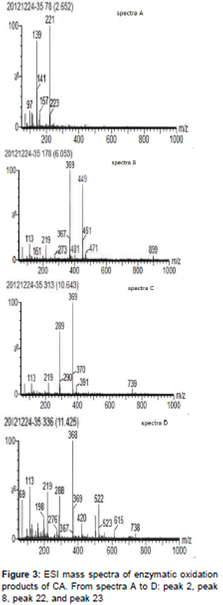
The total ion current chromatograms for peaks 2, 8, 22, and 23 generated under the same conditions after a 10 min reaction revealed obvious increases, suggesting that the substances from peaks 2, 8, 22, and 23 were products of the enzymatic oxidation of CA. In addition, peak 9 was CA and revealed a decrease in CA content as the duration of the reaction was extended. Conversely, the substances from peaks 8, 22, and 23 exhibited obvious increases. The mass spectral analysis of these substances is shown in .
The retention times of the CA enzymatic oxidation products in peaks 2, 8, and 22 were 2.65, 6.05, and 10.64 min, respectively. The abundant ion with m/z 369 in the MS spectrum corresponding to peak 22 indicated that the peak represented hydroxyl CA (molecular mass 370) with the loss of an H+ ion due to ionization. In addition, the approximately double molecular mass of the m/z 739 ion suggested that the substance from this peak was the monomer of hydroxyl CA. The m/z peaks in the spectrum corresponding to peak 8 (6.05 min retention time) revealed similarities to the m/z in the spectrum corresponding to peak 22. Consequently, we concluded that the products represented by peaks 8 and 22 were hydroxyl derivatives of CA. The structural characteristics of peak 2 were difficult to discern due to its small molecular mass, which indicated that it might not be an oxidation product of CA. In addition, due to the existence of characteristic ions with m/z 368 and 738 in the mass spectrum corresponding to peak 23, this fraction was believed to be p-quinone of CA because the peak with m/z 738 was nearly double that of m/z 368.
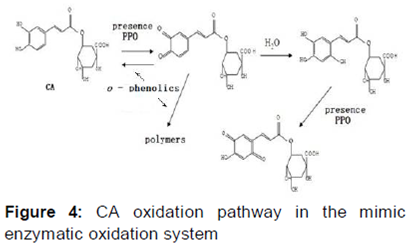
Oxidation pathway of CA in honeysuckle
Based on the results of the LC-MS analysis of the CA oxidation model system, we proposed a schematic diagram to describe the CA oxidation pathway ().
Discussion
In the simulated enzymatic oxidation system, hydroxyl CA, p-quinone derivatives, and o-quinone derivatives of CA were detected, indicating that CA generates hydroxyl CA in an enzymatic oxidation system. However, the o-quinone monomer of CA was also detected, which was most likely due to its instability and ready participation in the oxidation reaction and production of o-quinone and p-quinone CA derivatives. Nonetheless, despite the detection of the o-quinone intermediate monomer reaction during the enzymatic oxidation, the oxidation process and CA enzymatic oxidation reaction were consistent.
Conclusion
In the simulated enzymatic oxidation system, hydroxyl CA, p-quinone derivatives, and o-quinone derivatives of CA were detected, indicating that CA generates hydroxyl CA in an enzymatic oxidation system. However, the o-quinone monomer of CA was also detected, which was most likely due to its instability and ready participation in the oxidation reaction and production of o-quinone and p-quinone CA derivatives. Nonetheless, despite the detection of the o-quinone intermediate monomer reaction during the enzymatic oxidation, the oxidation process and CA enzymatic oxidation reaction were consistent.
Declarations
Acknowledgement
References
Archives


News Updates